Article 5: Motor Proteins, an Engineering Marvel

Finding the protein in the haystack!
Darwinian Evolution literature talks a lot about the creation of new proteins due to random mutations! We have talked previously about the ridiculous probabilities of finding a functional protein from the available space of possibilities in a typical protein peptide chain of 150 amino acids! Such probability can be as small as 1 in 10^77.
To appreciate this, we need to consider that this is in the order of finding a specific right atom that has gone in the whole universe, by pure chance! (We can estimate the number of atoms in the universe to be in the range of (10^78) – (10^82) atoms)!
Ref: Estimation of functionally beneficial proteins: https://www.sciencedirect.com/science/article/abs/pii/S0022283604007624 .
In this session, we will start to appreciate how those molecules can produce very complex functions on the molecular level, where those functions do not just arise from the development of a new protein molecule, but even from the 3D shape to which this molecule folds, and from the interaction of this molecule with other proteins and components to form systems that deliver specific functions in a way that is quite inconceivable to be the product of a random process! (Read about proteins here on nature education: Protein Structure: https://www.nature.com/scitable/topicpage/protein-structure-14122136/ ). It is also interesting to note that there are 500 amino acids in nature, out of which, specific 20-22 amino acids are used in living organisms, 20 of which are transcribed and translated from the genetic system
Ref: 20 Amino Acids that Make Up Proteins: https://www.ajinomoto.com/aboutus/amino-acids/20-amino-acids .

The Distribution Problem
Imagine a city where materials are imported from other cities, and goods produced at different parts of the city, and consumed at other parts, and even some goods are exported to other cities. An obvious essential solution is one of transportation and distribution of the materials and goods through the city. It occurs that the living cell is a metropolis where countless products are produced and moves around the cell to the right places where they are needed. Not only products, but even the very factories that produce those products can be mobile and get moved around the cell to go to another place.
It is striking to know that the cell has those robots that move around distributing materials and products around the cell; they are called “Motor Proteins” (see Session 5, and Supplementary Material section for videos about Motor Proteins).
Yet, a transportation system also means roads and railroads, a traffic system, and even a traffic law! Can we expect to find all of this in the cell, or equivalents to it?! Well, amazingly enough, those motors work together with special “roads” in the cell that are called Microtubules (tubulin), and Microfilaments (actin)!

How to detect intelligent System Design?
We recognize that a system is designed by an intelligent agent, when we see evidence of intelligence, in such a way, we can make a distinction between what is for example a rock that is carved by the elements, versus a status carved by an artist, or some metal scarps sitting together from a metal coffee making machine! In the case of the coffee machine, the metal pieces are brought together having specific shapes and dimensions, with specific interactions among them, so that the input material (the coffee and water and the heating) will produce the required outcome (the coffee) according to the design!
So, designed systems are ones where various parts are put together in a specific way, such that the overall system performs a certain function. So, when we see parts that have been put together, and they integrate and interact with each other so they complement each other to the fulfilment of the required function.
Motor Proteins

Imagine a little nanobot that moves in the inside of a cell, carrying a vesicle produced at one part of the cell to another part of the cell where it is needed! This bot has two legs that it uses them to walk, has an energy consumption mechanism that it uses to energize those legs, and at the top of its body there is hands that grab to the cargo to carry it around! This looks like science fiction, but it is not. It is what the three families of motor proteins do; myosin, kinesin, and dynein!
Those motor proteins have a body, two legs, and hands to carry the cargo.
Kinesins for example, one family of motor proteins, has a body made of two coiled protein spindles, attached to it the legs with have two feet aka “motor head domains” (which attach to the microtubules) attached to the body through two neck linkers which. On the cargo side, there will be a protein tail that would have a protein adaptor that is cargo specific to which cargo is attached, aka “cargo binding tail domains”.

Cargo-specific motor design
Interestingly enough, you will find a protein motor with specialized cargo tails that are specific to bind to the specific types of cargo!
The specificity of the design raises a question, those motors are essential for the transportation of this cargo in eukaryotic cells, so, if evolution is right, how did eukaryotes function without the presence of those sophisticated locomotives, and how do such a system develop with such specificity?! Can this be produced by a random mutation process?
Read this article in nature to appreciate the complexity of one of the motor protein families (Kinesin) and its various specialties in the cell (Kinesin superfamily motor proteins and intracellular transport: https://www.nature.com/articles/nrm2774?proof=t).

Microtubules, railroads of the cell
The motor proteins use a system of “roads” and “Scaffolding” in the cell that it can navigate, collectively can also be called the cytoskeleton, which has actin microfilaments and tubulin microtubules (MT). (Read about microtubules here: https://en.wikipedia.org/wiki/Microtubule )
It should be noted that MT has polarity which produces directionality, i.e. it has a positive and a negative end. MT tubulin has two proteins, an alpha and a beta tubulin, that attach in sequence typically forming 13 protofilaments that get arranged side by side to form the hollow Microtubule. Tubulin tends to have its negative end towards the cell center, while the positive end is towards the periphery.
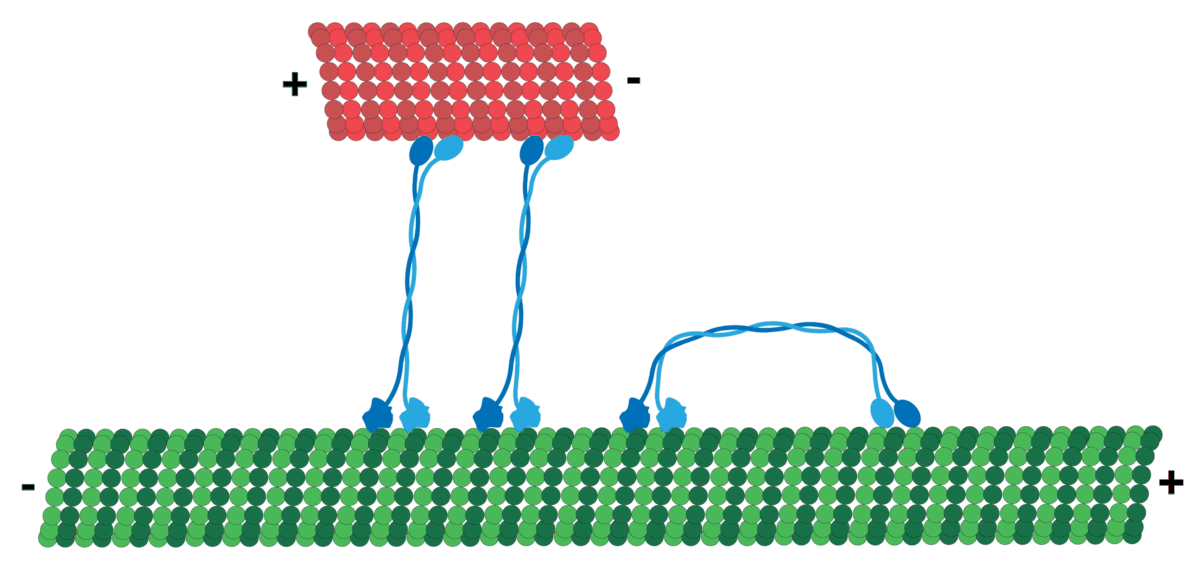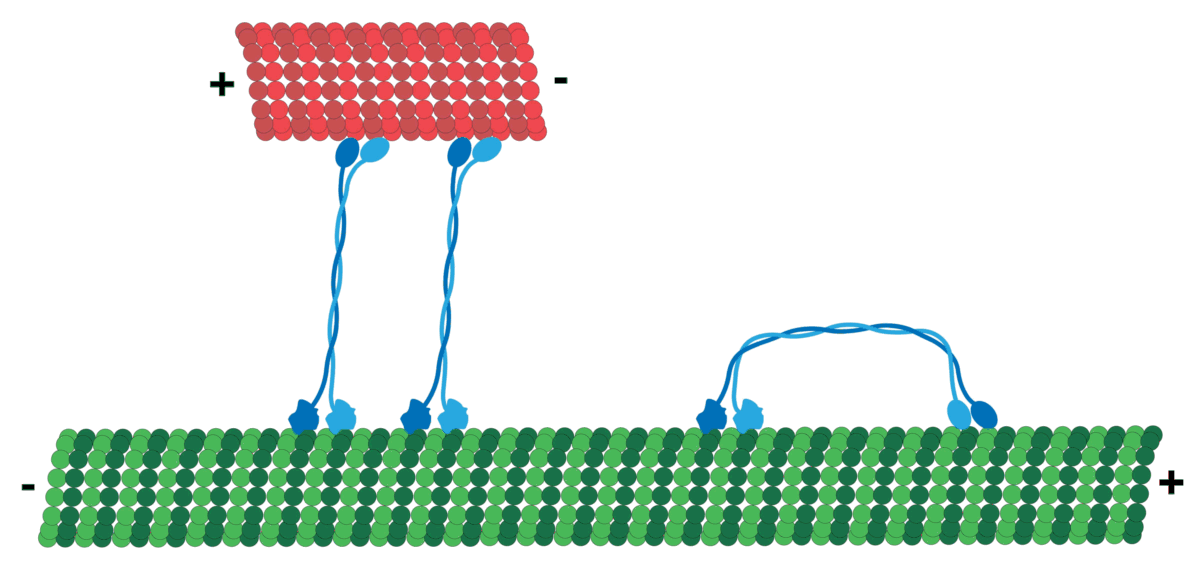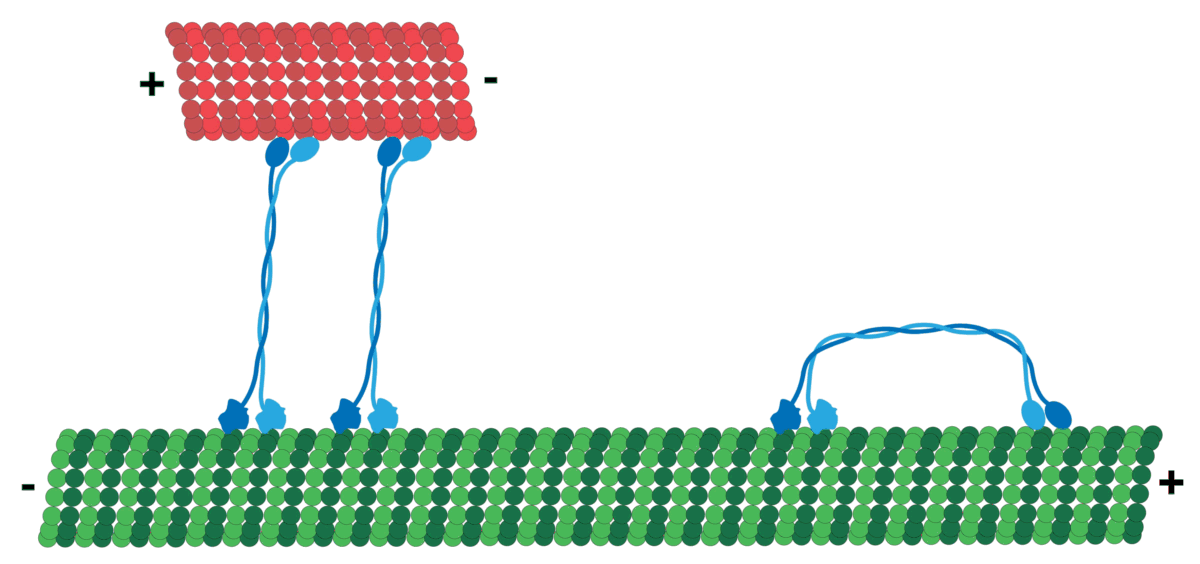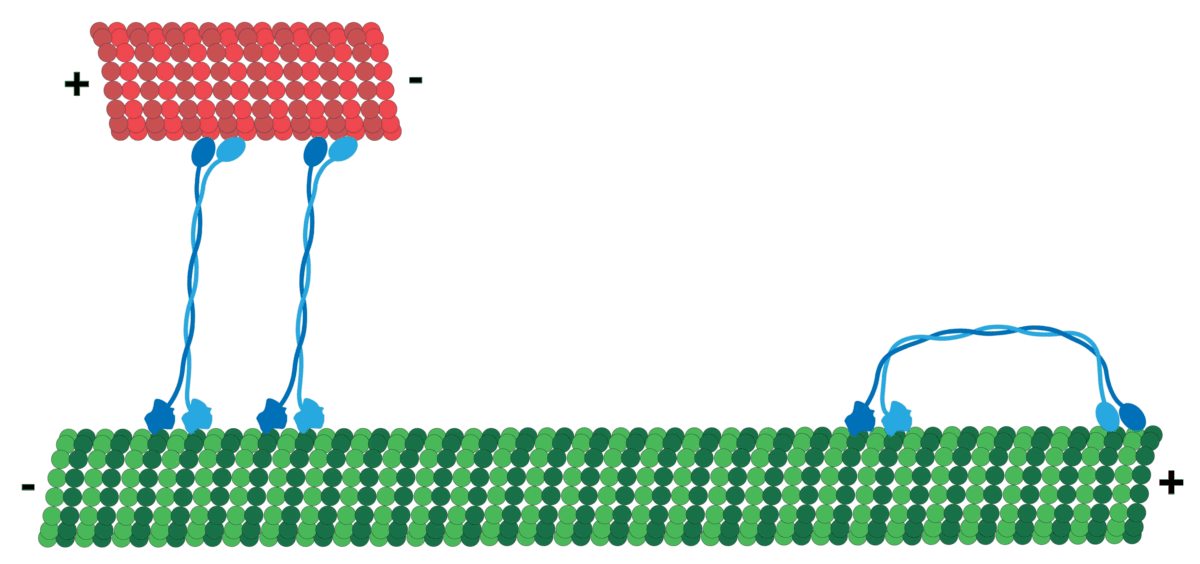
The motors generally navigate in a specific direction on the roads determined by polarity! So, cargo that should go, for example, from the inner part of the cell to the cell membrane will be loaded on a certain motor (e.g. Kinesis motor), and cargo moving the other way around can use a different one (e.g. Dynein motor)!
However, recent studies uncovered an unexpected result that some motors can move in the opposite direction that it typically uses (motor kinesin protein KlpA). (Read: Scientists discover a molecular motor has a ‘gear’ for directional switching: https://phys.org/news/2017-01-scientists-molecular-motor-gear.html
They discovered that KlpA is able to move in opposite directions – an unusual finding. KlpA-like motor proteins are thought to be exclusively one-way vehicles. The researchers also discovered that KlpA contains a gear-like component that enables it to switch direction of movement.

Motor feet (also known as motor head domains, which are connected to the motor body or spindle with a neck linker) bind to the microtubules attaching the motor to its road, yet a series of four chemical reactions would separate one leg from the microtubule, move it forward, attaches it again, and then separates the other leg, causing the walking effect of the motor. The reactions are energized and catalyzed through the consumption of energy stored in ATP molecules!
Compatibility at its best!
Once again, our attention is grabbed by the integrated nature of the design of the motor proteins, on the one hand its compatibility with the microtubules to be able to attach to them, and on the other hand with an ADP-ATP cycle that will accurately detach one leg, move it forward, then reattach the leg to the microtubule, and then the cycle repeats with the other leg, converting chemical energy into kinetic energy that generates a force driving the motor and its cargo down the rail road which is the microtubule! (Watch a very interesting video explaining the catalytic process here: https://youtu.be/YAva4g3Pk6k ).

Roads on Demand!
Another amazing fact is the presence, in the first place, of the microtubule (MT) (the “rail roads”) from the required origin of cargo to the required destination. It turns out that the microtubules can specifically assemble to produce the road for delivery of a certain cargo, and then disassemble when there is no further need for it. An example for this is the process of division of cells, where microtubules would rearrange into two centrosomes with radiating microtubules from them, and motors would take a critical signaling role, and then upon the end of the division process, the structure is disintegrated.
It should be interesting to read this article to notice how we, humans, are reverse engineering this brilliant transport system to unlock now horizons in nanotechnology and synthetic chemistry (https://arstechnica.com/science/2013/11/with-dna-and-proteins-researchers-build-transit-for-lugging-molecular-cargo/).
 To add to the marvels of this system, it was found that in some cases (Eg5 motor with MT), the very attachment of the kinesin to the MT will catalyze the MT to straighten up and elongate; so we are looking at a system where a vehicle that wants to go to a specific destination will get the road built for it until it arrives, and then maybe it will disassemble when there is no further need for it! (Read: Kinesin-5 Promotes Microtubule Nucleation and Assembly by Stabilizing a Lattice-Competent Conformation of Tubulin: https://www.sciencedirect.com/science/article/pii/S096098221930689X):
To add to the marvels of this system, it was found that in some cases (Eg5 motor with MT), the very attachment of the kinesin to the MT will catalyze the MT to straighten up and elongate; so we are looking at a system where a vehicle that wants to go to a specific destination will get the road built for it until it arrives, and then maybe it will disassemble when there is no further need for it! (Read: Kinesin-5 Promotes Microtubule Nucleation and Assembly by Stabilizing a Lattice-Competent Conformation of Tubulin: https://www.sciencedirect.com/science/article/pii/S096098221930689X):
Eg5 motor domain is sufficient to promote microtubule nucleation and assembly. Eg5 binding to tubulin promotes curved-to-straight transition. Eg5 binding and microtubule assembly can be treated as coupled equilibria. Loop11/α4 junction is necessary and sufficient for polymerase activity.
Is it obvious enough that this is a super integrated and harmonized system design?!
Now, with this super integration of this system of motor proteins, the tracks they use, the compatibility with their cargo, and the ATP-ADP cycles that cause them to stick and move, is it at all possible that this system came about through “random” mutations and natural selection?!!
I find the following video specifically offering a fascinating graphical representation of the walking kinesin: The Workhorse of the Cell: Kinesin: https://youtu.be/gbycQf1TbM0

Traffic lights, Sign Posts, and Traffic Law?!!
A recent amazing study uncovered a shocking mechanism, where a specific type of kinesin motors is guided to ferry chromosomal motion during mitosis (a stage in cell division), by specific markers on microtubules that act like sign posts, so that the kinesins will cause the chromosomes to be properly positioned and aligned in preparation for division! Researchers essentially tested this by removing the markers (which are special modifications on tubulin) and noticed that guidance process failed! Read from https://phys.org/news/2015-05-cell-division-chromosomes-microtubule-highway.html and Quote:
The Grishchuk lab showed that transport by a molecular motor protein called CENP-E kinesin is strongly enhanced on certain microtubule tracks. “This biophysical study was complemented by cell biological experiments carried out by the Maiato lab in Portugal, leading to the discovery that CENP-E-kinesin motors failed to take chromosomes to their alignment position if the guiding marks were eliminated”
Very specific design once again!
Now, we have to think once more, whether this kind of system, that is not only clearly designed with integration, efficiency, and compatibility in mind; but we are struck by “guidance” and external information to the system that is implanted in the system to guide the execution of the process in real time. This is like looking at a car that comes from the manufacturer with an integrated map of the city of operation to guide its navigation in cooperation with GPS information.
Is it even possible to consider that this has come about by random mutation?

The amazing kinetochore
Having spoke about guiding chromosomal alignment during mitosis, it is mandatory to talk about the kinetochore! One of the most complex structures in cell biology! It is an engineering marvel that is an interface between chromatin (packaged chromosome filaments) and the cytoskeleton that should pull chromosome pairs apart so that each of the new daughter set will get its share of the chromosome pairs. It needs to do it in the right time, with the right synchronization, when the whole cell is ready and each cell has what it needs. It is a mechanical tension, sensing, and signaling system, where the kinetochore interacts with the microtubules until (in a mysterious way) it senses that the time is right, and then you will see protein motors carrying a stop signal away from the kinetochore which is the final GO traffic light so that the system will pull the chromatids apart and the whole thing acts simultaneously to create the new daughter cells.
This can only be described in images rather than in words, and this video is the best that I can propose to you to watch (watch : Kinetochore and Mitosis: https://youtu.be/IvJrDsRuWxQ).
We cannot be here without protein motors!!
if you have watched the video and read well the above material, you will realize that eukaryotic life (all animals and plants) is largely dependent on those proteins to exist!
So, we need to notice that prokaryotic life (mainly bacteria) did not have those protein motors (though there is evidence it has simple cytoskeleton filaments in them). So, where did those complex and marvelous “vehicles” comes from to be that prevalent and abundant with all those diverse types that are cargo specific in eukaryotic life?
Eukaryotic life needs the motors, and the motors are made by translation of eukaryotic DNA… does that seem like a paradox?
Motors contract your muscles and make you hear!
We can go on and on mentioning the marvels that motor proteins can do, but one more thing that makes our very muscle move us around is the interaction of myosin motor proteins with actin filaments in our very muscle. The action depends on myosin heads binding to the actin and then pulling it in the contraction direction when the nerve signal causes release of calcium ions in the muscle. (watch: Excitation-Contraction Coupling: https://youtu.be/LlgaziPCFU0, and watch: Muscle Contraction – Cross Bridge Cycle, Animation: https://youtu.be/BVcgO4p88AA).
After seeing those two videos, it becomes very clear that
Motors causing you to hear!
If you want to be further amazed at what a protein motor can do, here is something about one that converts voltage to force and has piezoelectric properties, and is used in the cochlear cavity of mammals to make hearing possible! (https://en.wikipedia.org/wiki/Prestin).
Origins of Motor Proteins
Motor proteins are prevalent in Eukaryote (of which are animals and plants), but such complex motor proteins are not present in prokaryotes at a comparable level of complexity and diversity. Research shows that motor proteins can be traced to a hypothetical Last Eukaryotic Common Ancestor (LECA), but not easily prior to that!
So, where did those protein motors come from? Are they really possible to be there by “random mutations” and natural selection after all what we have witnessed? There are tens upon tens upon hundreds of those motors, configured and matched to numerous functions and cargo, and performing a very diverse set of functions without which eukaryotic life as we know it cannot exist.
Is it fair to assert (as the ToE obviously does) that such complexity and very obvious integrated system design came about by mere chance through “random” mutation and natural selection?!! (This paper discussed this topic for further reading: Patterns of kinesin evolution reveal a complex ancestral eukaryote with a multifunctional cytoskeleton: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2867816/). And the question remains practically unanswered, marking another paradox to the theory of evolution!
Motor Protein Paradoxes
How can Eukaryotes be a product of prokaryotes with all this innovation of motor proteins in Eukaryotes?!
Eukaryotic life is dependent on motor proteins, which are produced by transcribing and translating eukaryotic DNA, which is on eukaryotic chromosomes that can only exist if motor proteins are there in the first place to support eukaryotic life including eukaryote cell division. However, motor proteins (e.g. Kinesin and Dynein) are not present in prokaryotes! Since every eukaryote that this engagement between motors and cell division, it is very challenging to imagine how eukaryotes can come to existence from “evolutionary” perspective considering this one aspect (apart from many other perspectives).
Integration of Motors, Microtubules, ADP-ATP cycle!
Motors cannot function without microtubules, and microtubules (in terms of serving transportation) will not function without motors, so, if motors or microtubules evolve without the other, it will be deemed useless and eliminated by natural selection.
Moreover, ATP as a source of chemical energy is ancient in life, since prokaryotes (Ref: 15367437113476 1..30 (elifesciences.org)), so any locomotive system that evolves that uses energy other than ATP would have been eliminated.
Hence, we need an evolutionary system that seeks and finds the components of the motors with their enormous complexity, and binds them together to form the walker, given the microtubules will simultaneously evolve, such that their composition will match the heads (feet) of the motor and their neck-linkers, and both of them are fully matched with ATP-ADP hydrolysis to convert chemical energy to kinetic energy exactly in the manner that will choreograph the walking motion!
The simultaneous existence of all system components is paradoxical.
Motor specificity to a variety of cargo!
Cargo types have corresponding motor types that have the matching adapters to connect to them. Cargo can be large organelles or vesicles, like a mitochondrion, lysosome, or a liposome. Without the specific motor, the cargo will not move, causing cellular dysfunction and death.
Motors have adaptors that are specific to cargo types, so the right cargo will be picked up by the right motor from the right place to the right destination!
So, if the organelles/cargo existed before the motors, they will not deliver proper function, and if the motors existed prior to the cargo, the motor will be eliminated being deemed useless.
Another chicken and egg paradox.
Randomness or design?
Is it more reasonable to infer that such an integrated system is the product of systems design, or just chance through random mutation!
I leave you to contemplate on those questions, and ask yourself, if it is possible for randomness to generate such a sophisticated, highly specific intricate system!
Ahmed Abd ElSattar
Finding Truth